Five digits radiating from a palm, an arrangement both flexible and strong—capable of playing a piano, wielding a hammer, offering a comforting touch. The hand is our most familiar body part, central to most everyday tasks, from dressing and driving to cooking and texting. Yet from an evolutionary standpoint, it remains largely mysterious, particularly when it comes to the earliest stage of its origin. Other four-limbed creatures—tetrapods, as they are known—have hands that look and function quite differently than ours do. In birds and bats, they help to form delicate wings; in elephants, they support limbs as big around as tree trunks. But the basic structure is the same. In 1859 Charles Darwin remarked on the similarities in On the Origin of Species: “What can be more curious than that the hand of a man, formed for grasping, that of a mole for digging, the leg of the horse, the paddle of the porpoise, and the wing of the bat, should all be constructed on the same pattern, and should include the same bones, in the same relative positions?”
Darwin proposed an elegant explanation: these diverse animals share this pattern because they evolved from a common ancestor that possessed limbs with digits. In the more than 160 years since Darwin advanced his revolutionary idea, evolutionary biologists have marshaled evidence from paleontology, genetics and embryology that has proved him right. Their efforts have illuminated the shared ancestry of tetrapods, which evolved from fish; shown that the bones that make up the human hand are also found in frogs and birds and whales; and identified some of the genes that control the development of hands and wings and flippers, among other variations. But the first chapter of that story—the bit where the hand and wrist evolved from bones in the fin of an ancestral fish—has remained murky at best because scientists have lacked sufficiently complete fossils of transitional creatures between fully aquatic fish and land-roving tetrapods.
This past March we unveiled an extraordinary fossil—a complete skeleton of a 375-million-year-old fish, Elpistostege watsoni—that goes a long way toward filling that gap in understanding. The fossil preserves in its fins bones comparable to the ones that make up our fingers, showing that digits evolved before vertebrates left the water. This discovery overturns the conventional wisdom about when and how the hand evolved and shines new light on the rise of tetrapods, a pivotal event in the history of life on earth.
Murky Origins
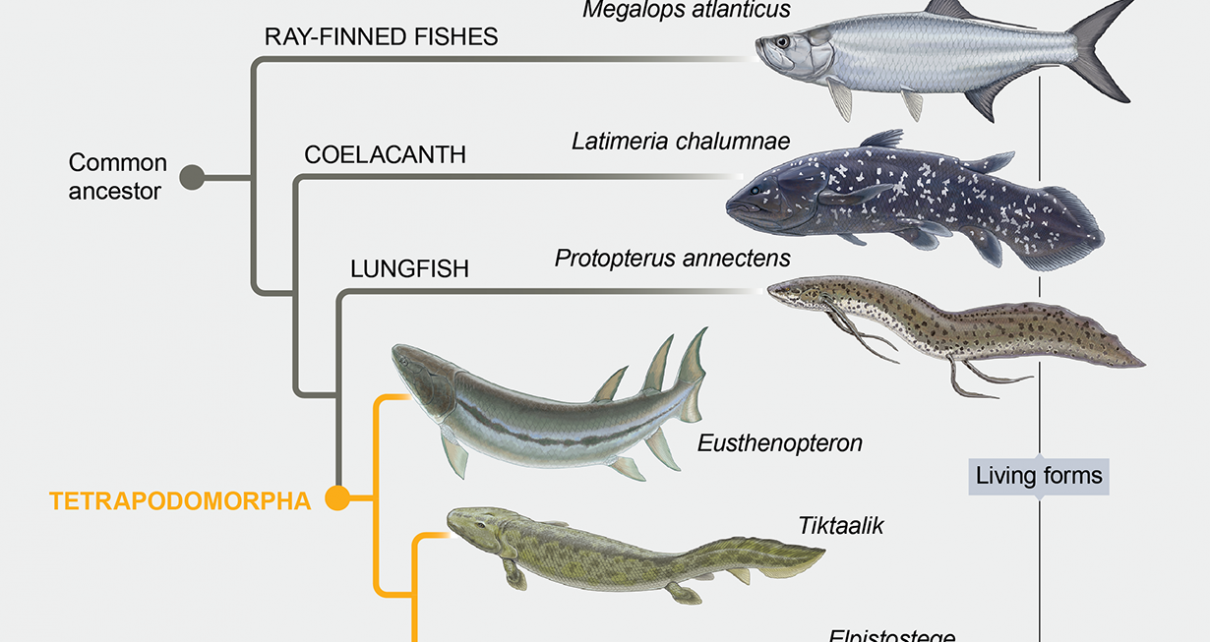
Until recently, scientists’ grasp of the evolutionary transition between fishes and early tetrapods hinged mainly on several spectacular fossils that seem to bridge these two groups. One is from a fish called Panderichthys rhombolepis from the Baltic region and dates to the Middle to Late Devonian period (around 384 million to 379 million years ago). With its elongated humerus and large radius and ulna (the upper arm bone and forearm bones, respectively, in tetrapods) and its tetrapodlike skull bone pattern, Panderichthys offered the first clues that the group of fishes to which it belongs were the closest fishes to the tetrapods. The group is called the elpistostegalians, after the then poorly known Elpistostege from eastern Canada.
In 2006 Neil Shubin of the University of Chicago and his colleagues announced the discovery of another elpistostegalian fish fossil, 380-million-year-old Tiktaalik roseae from the Canadian Arctic. Tiktaalik was a real game changer in revealing a slew of new data showing that the pectoral fin was highly advanced in these fishes—more so than any other known fossil—with both well-developed arm bones and mobile wrist joints. Its skull also had distinctive features, including a long, flat snout and a specialized braincase—traits shared by tetrapods.
Together, this and the other known elpistostegalian fish fossils suggested that a number of hallmark traits of tetrapods originated in their piscine predecessors, including land-worthy arm bones and joints. But what these fishes did not appear to have were fingers. In the case of Panderichthys, bony elements that many researchers initially thought were rudimentary digit bones were later rejected as such. And the Tiktaalik fossil, for its part, did not preserve the complete tip of the pectoral fin, where one would expect to find digit bones if the animal had them. The available evidence left experts to conclude that fingers were not part of the initial fin-to-limb transition. Instead they appeared to have evolved later, after tetrapods had already staked a claim on terra firma.
In science, however, knowledge is not written in stone. It is subject to change in light of fresh evidence. New discoveries can necessitate revision of the textbooks. Our recently described Elpistostege fossil, which was unearthed in 2010 at the UNESCO World Heritage Site of Miguasha in Quebec, is one such find. It is not a new species of elpistostegalian. Rather it is the original founding member of the band. But this time we have a complete, perfect specimen. And it has led us to propose a different theory of how fingers evolved and gave rise to the vertebrate hand structure that persists in the more than 33,800 species of tetrapods alive today, including humans.
To appreciate the role of Elpistostege in shifting our perspective on how hands evolved, it helps to know a bit about the history of its discovery. In the summer of 1937 two young British paleontologists were scouring the cliffs of Chaleur Bay along the southern shore of the windswept Gaspé Peninsula in eastern Canada. Thomas Stanley Westoll and William Graham-Smith were looking for Devonian-age fossils, and the cliffs were known to be an El Dorado for such treasures. Local collectors assisted the paleontologists in making their discoveries and sometimes sold them fossils. One of the fossils that Westoll purchased from them was a small, fragmentary skull roof that was to become a cornerstone in our understanding of the evolutionary transition between fishes and tetrapods.

Back in Westoll and Graham-Smith’s day, scholars already suspected that tetrapods had evolved from the so-called lobe-finned fishes—creatures with fleshy, powerful fins, a group whose living representatives include the coelacanth and the lungfish. But they lacked fossils with intermediate anatomy to bolster the connection. Westoll’s skull roof seemed like something that might help fill the gap. Given the pattern of its skull bones, Westoll thought it could be the long-sought-after skull of a primitive Devonian amphibian. He named this unique specimen Elpistostege watsoni, from the Greek for “hoped for” and “roof.” In a brief paper published in 1938 in Nature, Westoll argued that the fossil provided “a perfect transition” between lobe-finned fishes and early four-legged animals.
Based on only a piece of skull, Westoll’s argument was met with skepticism. More of the animal was needed. But although paleontologists came from all over Europe and America to collect fossils from the cliffs of Chaleur Bay, an area now designated Miguasha National Park, no one collected a new specimen of Elpistostege.
Some 30 years after Westoll’s publication, however, serendipity intervened. Canadian fossil collector Allan Parent discovered the snout of an incomplete skull in the Miguasha Cliffs. It remained hidden in his private collection until his tragic early death. But in 1983 Parent’s brother brought the snout to the attention of the Miguasha park director, Marius Arsenault, who in turn enlisted Hans-Peter Schultze, an eminent specialist on fossil fishes from the University of Kansas, to identify this unusual specimen. Schultze immediately realized its importance: similarities in the arrangement and shape of the cranial bones preserved in this fossil and Westoll’s skull roof showed they belonged to the same species.
With more of the creature’s anatomy revealed, ideas about its identity began to shift. The features visible on the new snout suggested that Elpistostege was not an amphibian but a highly advanced lobe-finned fish. In their 1985 paper describing the snout, Schultze and Arsenault specifically suggested that it was closely related to the Baltic fossil fish Panderichthys. They also discussed another enigmatic specimen, a fossil containing a patch of scales and some vertebrae that Miguasha chief biologist Marc Brassard and one of us (Cloutier) had found a couple of years earlier in the same cliffs. Considering the similarities in the surface textures of the scales preserved in this fossil and the snout, Schultze and Arsenault proposed that it, too, belonged to Elpistostege.
The attribution of the fossil containing the scales and vertebrae to Elpistostege was important. In addition to being the only material then known from below the creature’s head, it had something the other two Elpistostege fossils did not: detailed provenance. The skull roof and snout were known to have been collected from the cliffs, but no one knew which rock layer they came from. In contrast, Brassard and Cloutier had recorded the exact stratigraphic location of the fossil they found: 90 meters above the lowermost layers of a distinctive geologic unit known as the Escuminac Formation.
In the years that followed, Cloutier and his collaborators returned to this part of the Escuminac Formation again and again to search for more remains of Elpistostege—to no avail. Still, the geologic and fossil evidence they did find allowed them to begin to reconstruct the animal’s environment, a channel that fed into an estuary. A picture began to emerge of Elpistostege as the largest fish—and probably therefore the top predator—in the waters it shared with some 20 other species of fishes.
Eventually that paleoecological data would gain new relevance. Late in the afternoon of August 4, 2010, while on patrol in Miguasha, park warden and naturalist Benoît Cantin found an unusual piece of fossilized fish tail embedded in the lower Escuminac rock layers on the beach at the foot of the cliffs, less than 250 meters from the park’s museum. The next morning Cantin, accompanied by two naturalist guides, unearthed the rest of the animal. It is the largest fossil ever found in the Escuminac Formation and arguably the most important one—a 1.57-meter-long complete skeleton of Elpistostege.
Seeing through Stone
Within days of Cantin’s finding the fossil, Cloutier was invited to study it. Because most of the skeleton was still entombed in rock, the first order of business was to get a better look at it by using computed tomography (CT). Cloutier recruited Isabelle Béchard, one of his former master’s students in paleontology, to help him image the specimen using a nonmedical CT scanner at the Institut National de la Recherche Scientifique (INRS) in Québec City. The scan showed that the specimen was absolutely complete—every single bone was preserved—but the resolution was not good enough to reveal the internal structure of the bones, which they wanted to be able to study. They had to find another CT scanner that provided better resolution. They decided that Elpistostege was going to travel some 3,900 kilometers south to the High-Resolution X-ray Computed Tomography Facility at the University of Texas at Austin, where each piece of the fossil would be scanned with the greatest precision available at the time.
Once the scanning was done, Cloutier and his colleagues began the meticulous work of preparing the fossil—removing the surrounding rock bit by bit to expose the bones within. They performed this work on a computer model of the fossil based on the CT scans as well as on the actual fossil. After a few months the body and the skull emerged from the rock, both virtually and in reality. The whole specimen was incredible, but the pectoral fins generated particular excitement because no one had ever seen a complete pectoral fin of an elpistostegalian before. Numerous skeletal elements were visible within the fins, surrounded by scales and fin rays. At first glance Elpistostege‘s fin looked quite similar to that of Tiktaalik, but Elpistostege‘s appeared to have additional bones in it. What were they?
In the years that followed, Cloutier and Béchard presented preliminary results of their analyses of the Elpistostege skeleton to colleagues at professional meetings. It was after one such presentation, in 2014 at the Society of Vertebrate Paleontology in Berlin, that Cloutier and one of us (Long) agreed over a beer to collaborate on the study of this remarkable fossil.
Long had been working on Devonian fish fossils from the Gogo Formation in Western Australia for many years. The skeletons of these ancient fishes are perfectly preserved in three dimensions, and some specimens show exceptional preservation of soft tissues as well. To study these fossils, which are found inside nodules of limestone, Long and his colleagues traditionally placed them in acid baths to dissolve the rock. More recently, they have shifted to imaging the fossils using CT and synchrotron scanners and sophisticated software to elucidate their fine anatomical structures before the acid baths, so that they can capture any soft-tissue preservation before the acid destroys it. The new Elpistostege specimen, with its exquisite preservation, seemed like a prime candidate for Long’s approach to imaging and digital preparation.
In 2014 Long visited Cloutier’s lab in Quebec and began working with his team on how to use different methods to process the imaging data from this fish. It took some trial and error with different types of data and software, but eventually we hit on a winning combination that would allow us to digitally isolate and study each bone. Alice Clement of Flinders University in Adelaide and Roxanne Noël and Vincent Roy, then master’s students working under Cloutier, went on to carry out this work.
When Clement eventually began segmenting out the pectoral fin, we were at the edges of our seats. As the first complete elpistostegalian pectoral fin ever discovered, it was certain to contain vital clues to the transition from fins to limbs. The preliminary results did not disappoint. They not only confirmed Béchard’s initial CT results suggesting the presence of extra bones in Elpistostege‘s fin but also showed those bones in far greater detail. Now we could see that the fossil included an unexpected series of many small, tightly packed bones. Typically the end of the pectoral fin skeleton contains small bones called radials that support the rodlike fin rays. The bones evident in this part of Elpistostege‘s fin were in the right place to be radials, but the large number of bones and the way some of them were arranged in discrete rows suggested that they were something else. We strongly suspected that these never-before-seen bones hidden in the pectoral fin of this ancient fish were actually digit bones similar to the ones found in the fingers of tetrapods. We identified two digits that were each composed of multiple, articulated bones, as well as three possible digits each composed of a single bone.
Why is the case for digit bones in the pectoral fins of Elpistostege more compelling than the earlier argument for digit bones in the pectoral fins of Panderichthys? The putative digit bones in Panderichthys are irregular in shape, and none of them show articulation with other bones in the standard way digit bones, or phalanges, do in the human hand. We surmise from comparison with the bones of the Elpistostege fin that the mystery bones in the Panderichthys pectoral fin are probably equivalent to some of the carpal bones in the wrists of tetrapods.
Fish out of Water
As the team finished isolating the entire pectoral fin and girdle such that every bone could be studied in every aspect, we also began segmenting out internal features of the skull and braincase. This work would help inform our understanding not only of Elpistostege‘s anatomy but also of how it is related to other early fishes and to tetrapods.
To figure out where Elpistostege belongs in the family tree, we needed to compare it with other species, paying special attention to their shared distinctive features. Working with Mike Lee of Flinders, a leading expert on methods for ascertaining relations among species, we ran a phylogenetic analysis of 202 characteristics across 43 species. In the end, we were astounded to find that Elpistostege appears to be more closely related to the crown tetrapods—the group that includes all living tetrapods and their last common ancestor—than the well-known Tiktaalik is, though not by much. Our best guess is that the unique features of the pectoral fin, which is not fully preserved in Tiktaalik, plus some newly discovered features of the pectoral girdle (the bones that connect to the fins), pulled Elpistostege up a notch above Tiktaalik on the evolutionary ladder.
Combining what we see in the skeleton of Elpistostege with what the phylogenetic analysis tells us about its place in the family tree, we can reconstruct how the basic plan for the hands of tetrapods, including humans, originated. The presence of small rows of bones we identified as digits in the pectoral fin of Elpistostege shows that the arrangement first evolved in the fins of advanced lobe-finned fishes back at the start of the Late Devonian period, more than 380 million years ago. It is likely that they served to bear weight because having many tiny bones in alignment in this region would have given the outer part of the robust fin the flexibility needed to push the fish upward.
Why might a fish benefit from being able to maneuver in this way? The skull of Elpistostege contains a clue: at the back of the head is a pair of large holes called spiracles. Some modern-day fishes that breathe air have similarly large spiracles. For a long time, the function of these holes was uncertain. In a study published in 2014, Long worked with a team of ichthyologists at Scripps Institute of Oceanography in La Jolla, Calif., led by the late Jeff Graham, to nail it down. Analyzing the spiracles of the living bichir Polypterus, we showed that they are instrumental in breathing air. Assuming the spiracles served the same purpose in Elpistostege, the ability to employ the fins to do a push-up in the shallow rivers and estuary this fish inhabited—and thereby get the head out of water for a breath of fresh air—could have been advantageous.
Elpistostege was not necessarily restricted to the aquatic realm, however. Today’s lungfish and some catfishes can propel themselves along land for short periods with their fins. With its far more powerfully built fins, Elpistostege was probably that much more capable of venturing ashore.
Baby Steps
In addition to upending the received wisdom about when fins became limbs, our discovery of digit bones in Elpistostege bears on efforts to understand the genetic and developmental changes that powered this transformation. Just a few decades ago scientists interested in this question did not have much to go on. Back then, there were only a few examples of fossil fishes with pectoral anatomy transitional between fins and limbs, and they hinted only that the arm and forearm bones evolved gradually. In contrast, it looked as though tetrapods evolved the hand and wrist from the ancestral fish fin all at once. But was it actually possible for such drastic change to occur so abruptly? Or was the seemingly sudden origin of the hand and wrist simply an artifact of an incomplete fossil record?
In a milestone paper published in 1991, the late developmental biologist Peter Thorogood proposed that the hand and wrist really could have originated quite suddenly. He based his proposal on comparisons of embryological development in fishes and tetrapods. In early-stage embryos of both groups, a structure called the apical ectodermal ridge (AER) acts as a signaling center that guides development of the fin and limb, respectively. Located in the tip of the budding fin and limb, the AER secretes chemicals that promote the outgrowth of the underlying tissue. In fish, the AER is active for only a short while before it transforms into another signaling center, the apical ectodermal fold (AEF), which directs the formation of the fin rays. In tetrapods, however, the AER is much more active in directing development because it persists far longer in the embryo—it does not ever morph into the AEF; thus, the fin rays do not form. So it has more time to develop the other bones of the appendage, which are the basis for making a hand. Thorogood suggested that the loss of the AER-AEF transition—a relatively simple tweak in the grand scheme of things—could produce both the loss of the fin rays and other characteristics of the pectoral fins and the simultaneous gain of the bones needed to form a wrist and digits.
.png)
The discovery of Tiktaalik, the first fossil fish to show not just the fish equivalent of our arm bones and some of the proximal wrist bones (those closest to the center of the body) but also bones from the distal part of the fin corresponding to our distal wrist bones, spurred research into the genetic and developmental underpinnings of the fin-to-limb transition. Investigators were keen to figure out which bones in the ancestral fish fin evolved into wrist and hand bones and which genes were responsible for generating that new morphology. Members of the Hox family of genes, which are well known to direct the different regions of the embryo to develop into the head, tail, and so forth, seemed likely to be involved.
In 2007 Zerina Johanson of the Natural History Museum in London and her colleagues studied the activity of one of these genes, HoxD13, in one of the closest living relatives of tetrapods, the Australian lungfish. Previous research had shown that HoxD13 is active in the developing tetrapod limb when the wrists and digits form. Johanson’s team showed that the gene is also active during the development of the radial bones in the lungfish fin. But whereas in tetrapod limb development HoxD13 has two phases of activity—an early phase associated with the arm and forearm development and a late phase associated with wrist and digit development—it appears to have only one interval of activity in lungfish-fin development, corresponding to the second phase of the gene’s activity in the developing tetrapod limb. The work suggested that tetrapod digit bones evolved from the radial bones in the fish fin. But wrist and hand could not have originated as a package deal, as Thorogood proposed, because lungfish and other living and fossil lobe-finned fishes have radials or wrist bones without having digits. There had to have been at least two evolutionary events, one that produced digits and another that produced wrists.
Elpistostege further complicates the story. It suggests that, contrary to Johanson’s argument, the radials of lungfish and other lobe-finned fish are not all equivalent to digits. Instead only the most distal ones are homologous to digit bones; the proximal radials are homologous to the wrist bones and the long bones of the palm. What is more, Elpistostege reveals yet another step in the fin-limb transition. Because it preserves both wrist bones and digits, as well as fin rays, Elpistostege shows that loss of the fin rays must have been another, separate phase in the evolution of the hand.
The anatomical condition of the pectoral fins and digits we have described in Elpistostege suggests that we still need to fine-tune our understanding of the genetic and developmental mechanisms that drove their evolution. Nevertheless, with the entire skeleton preserved and many further studies of it underway, this specimen of Elpistostege seems destined to serve as a Rosetta Stone to solve the mystery of how limbs evolved from fins—and thus how vertebrates conquered land.